 |
 Как обустроить мансарду?  Как создать искусственный водоем?  Как наладить теплоизоляцию?  Как сделать стяжку пола?  Как выбрать теплый пол?  Зачем нужны фасадные системы?  Что может получиться из балкона? |
Главная страница » Энциклопедия строителя
содержание:
[стр.Введение] [стр.1] [стр.2] [стр.3] [стр.4] [стр.5] [стр.6] [стр.7] [стр.8] [стр.9] [стр.10] [стр.11] [стр.12] [стр.13] [стр.14] [стр.15] [стр.16] [стр.17] [стр.18] [стр.19]
страница - 10
Результаты III: позвоночные, предпочитающие жить в водной среде
Переход к постоянному обитанию на суше можно рассматривать как ключевой момент в эволюции позвоночных [Северцов, 1967; Шмальгаузен, 1968]. С информационной точки зрения наземная среда - сложный лабиринт с довольно высоким уровнем неопределенности в отношении, например, места нахождения пищи или опасности. Многочисленные препятствия, существующие в окружающей среде, усложняют планирование адекватного поведенческого ответа, если этот ответ исключительно базируется на текущих сенсорных впечатлениях. Таким образом, наземная среда - серьезное испытание для интеллектуальных (прогностических) способностей животного, поскольку принятие решения должно быть сделано в условии высокой неопределенности на основе текущей информации. Считается, что определенные морфологические и физиологические изменения в организации мозга позволили адаптироваться полностью к наземному образу жизни [Северцов, 1939, 1967]. Эта точка зрения согласуется с идеей, что ключ к успеху должен главным образом зависеть от ассоциативных и познавательных способностей животного, поскольку эти свойства определят скорость обработки информации и пластичность исполнительных и гомеостатических систем, которые лежат в основе поведенческой реорганизации [Гальперин, 1976; Леонтьев, 1981].
Однако многие виды позвоночных, включая млекопитающих, возвратились к водному образу жизни. Часто под давлением отбора возвращению в воду способствовали такие особенности организма как плохо развитые скелетно-мышечная, сердечно-сосудистая, дыхательная и пищеварительная системы [Яблоков et al., 1972; Кочеткова, 1973]. Роль когнитивных особенностей для выживания и адаптации к наземным условиям среды как фактора эволюции, как правило, не рассматривали, полагая, что структурные и функциональные характеристики мозга были результатом экологической адаптации [Никитенко, 1969; Grant, 1985; Shettleworth, 1998, 2000]. Однако палеоневрологические данные четко указывают на то, что реорганизация ЦНС у позвоночных предшествовала другим морфологическим изменениям, и некоторые виды с более сложными умственными способностями (по оценке больших размеров мозга и гирификации) вымерли, тогда как с менее сложными умственными способностями (меньший мозг без борозд и извилин) выжили [Dechaseaux, 1962; Кочеткова, 1973; Edinger, 1975].
Многие факторы могли быть причиной вторичноводной адаптации у позвоночных животных в процессе филогенеза. Возможно, что и интеллектуальные способности также участвовать в этом процессе, поскольку некоторые морфологические реорганизации мозга, возникающие в процессе адаптации к наземной среде вместе с другими анатомическими и физиологическими дисфункциями могли привести к тому, что некоторые виды позвоночных оказались неспособными выжить на суше. Чтобы проверить гипотезу о том, могли ли когнитивные особенности быть фактором, содействующим возврату в водную, более простую с информационной точки зрения, среду, мы провели исследование четырех видов водных позвоночных животных. Два из них, караси (Carassius auratus) и карпы (Cyprinus carpio) были первичноводными, а другие два вида, черепахи (Clemmys caspica) и дельфины (Tursiops truncatus) - вторичноводными. Эти животные содержались в группах до и после опытов, а в экспериментальной среде их тестировали индивидуально.
Первичноводные виды (Рыбы)
До испытания в нашем эксперименте все рыбы обучались в течение трех недель получать пищу (маленькие кусочки мяса) в ответ на дергание красной пластмассовой бусинки в аквариуме содержания. Лабиринт был построен в экспериментальном аквариуме (90 x 80 x 60
см) и состоял из сходной пространственной структуры, используемый для наземных млекопитающих, но с незначительными модификациями (рис. 12). Он был более сложен, так как было больше альтернативных проходов, поэтому коэффициент разветвления в лабиринте рыб был выше (32 отсека по сравнению с 25, используемыми для наземных млекопитающих) и имел большее количество маршрутных вариантов решения задачи. Опыты проводились ежедневно или через день. До опытов рыбы были лишены пищи в течение 22 часов. Время опыта (ТОП) составляло 20 мин в начале экспериментальных процедур и было постепенно увеличено до 60 мин (см. объяснение в тексте).
-^-Х
свободное поле
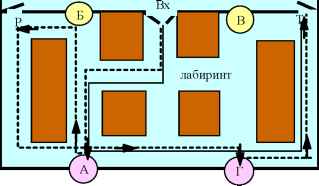
Рис. 12. Схема экспериментальной среды для рыб.
П - белая пластмассовая бусинка (1.4 см в диаметре) расположена в зоне свободного поля напротив входа (Вх) в лабиринт. Четыре бусинки меньших размеров (0.7 см в диаметре) находились в лабиринте, две из которых (А и Г) были связаны с получением пищи, а две других (Б и В) были ложными. Рыба должна была в условиях свободного выбора формировать оперантный 5-звенный навык: животное, помещенное в стартовое положение в свободном поле, должно было дернуть за бусинку П (звено 1) для того, чтобы войти в лабиринт (звено 2); войдя в лабиринт, рыба должна была приблизиться к бусинке-подкрепляющей кормушке А или Г, дернув за которую получала порцию пищи (звено 3); затем рыба могла посетить вторую подкрепляющую кормушку и получить вторую порцию пищи (звено 4), после чего животное должно было без какого-либо условного стимула покинуть лабиринт (через P или T) и выйти в свободное поле (звено 5) для того, чтобы иметь возможность многократно получать подкрепление при последующих заходах в лабиринт.
Приведенные в данной работе и более ранние исследования [Никольская, 1992, 1997], так же как другие работы [Лещова, Жуйков, 1989; Loppez et al., 1999, 2000], свидетельствовали о том, что рыбы способны формировать сложное поведение, если соответствующие экспериментальные условия были выполнены. В нашей поведенческой модели при ежедневной постановке опытов ни одна из двух изученных видов рыб не смогла самостоятельно решить предложенную задачу. В этих условиях после непродолжительного периода исследовательской активности в течение первых 3 опытов в поведении животных начинали возникать и нарастать невротико-подобные реакции в виде манежных движений, выпускания пузырей, бросковых ударов о дно аквариума или неподвижного зависания, и начиная с 7-го опыта, рыбы отказывались входить в лабиринт вообще. Эта двигательная депрессия (ЛА = 22.3 ± 10.5 зон в опыте; М. ± SD) сохранялась в течение последующих 20 опытов, и такое поведение (проявлявшееся как неподвижное зависание в воде) было неизменным. Увеличение
продолжительности опыта до 40, затем до 60 мин не привело к каким-либо позитивным изменениям.
Из-за отсутствия прогресса в обучении рыб мы увеличили межопытный интервал (МОИ) до 48 ч. Благодаря этому 30 % карасей и 27.3 % карпов смогли решить задачу и сформировать целенаправленное поведение, однако это произошло только при условии 60-минутной продолжительности опыта (рис. 13). Рыбы, отказавшиеся от решения предложенной задачи, проявляли признаки двигательной депрессии в течение всего времени опыта, независимо от его продолжительности. Переход на 48-ч МОИ у рыб, проявлявших более высокую двигательную активность как в аквариуме содержания, так и в лабиринтной среде, вызвал явное поведенческое оживление в виде многократных приближений к подкрепляемой бусинке и "маниакальное" ее дергание (6.7 ± 2.5 раза; М. ± SD). Однако при ТОП = 20 мин уже к 12 - 15-ой минуте нарастали пассивные (вертикальное ныряние с выпусканием воздушных пузырей и длительное зависание в воде) и активные (маятникообразные или круговые движения по стенке аквариума) стрессовые реакции. Начиная с 15 - 17-го опыта развивалась устойчивая двигательная депрессия, которая становилась преобладающей формой поведения рыб. Таким образом, опыты с ТОП = 20 мин при 48-ч МОИ оказались неадекватными для успешного решения предложенной задачи у изученных нами видов рыб (рис. 13).
60 50 40 30 20 10
г 0.7 0.6 - 0.5
0.4 0.3 0.2 0.1
0
8 I 12 40 мин
О п ы т ы
0
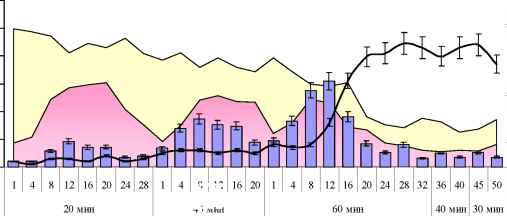
Рис. 13. Поведенческие показатели обучения рыб в зависимости от длительности опыта.
По вертикальной оси (слева) отложено число заходов в лабиринт (черная линия) и количество различных ошибочных действий: повторных подходов к подкрепляемым кормушкам и посещение ложных (голубые столбцы), по вертикальной оси (справа) - доля различных психо-эмоциональных проявлений, сопровождающих процесс обучения (желтая площадь) и доля активных невротико-подобных реакций (розовая площадь). Рисунок показывает, что динамика распознавания когнитивной задачи и формирование навыка у рыб происходило сходным с наземными млекопитающими образом (см. рис. 4В), если условия для протекания ассоциативных и интегративных процессов были выполнены.
Поведение рыб резко менялось как только ТОП было увеличена до 40 мин. Двигательная активность у обучавшихся рыб возрастала вдвое (ЛА = 98.5 зон/опыт и 210.3 зон/опыт при ТОП в 20 и 40 мин соответственно), а стрессовые проявления резко снижались (рис. 13). Исследовательская активность у рыб устойчиво сохранялась на протяжении всего времени опыта, возрастала маршрутная и семантическая вариабельность (рис. 14). Увеличение
содержание:
[стр.Введение] [стр.1] [стр.2] [стр.3] [стр.4] [стр.5] [стр.6] [стр.7] [стр.8] [стр.9] [стр.10] [стр.11] [стр.12] [стр.13] [стр.14] [стр.15] [стр.16] [стр.17] [стр.18] [стр.19]