 |
 Как обустроить мансарду?  Как создать искусственный водоем?  Как наладить теплоизоляцию?  Как сделать стяжку пола?  Как выбрать теплый пол?  Зачем нужны фасадные системы?  Что может получиться из балкона? |
Главная страница » Энциклопедия строителя
содержание:
[стр.Введение] [стр.1] [стр.2] [стр.3] [стр.4] [стр.5] [стр.6] [стр.7] [стр.8] [стр.9] [стр.10] [стр.11] [стр.12] [стр.13] [стр.14] [стр.15] [стр.16] [стр.17] [стр.18] [стр.19]
страница - 5
решения задачи. Следует обратить внимание на то, что на этапе стабилизации навыка данная группа крыс никогда не выполняла оптимальную структуру решения задачи с первой пробы в опыте.
Признаки извлечения целостного решения из памяти отсутствовали, вместо этого можно было наблюдать феномен воссоздания его по механизму принятия решения [Ещенко, 1998; Костенкова с соавт., 1999]. Опыт начинался с извлечения инстинктивной программы (Вход -Кормушка - Выход; см. рис. 1), нарастающее возбуждение из-за совершаемых ошибок приводило к возникновению в поведении фрагментов задачи, которые затем объединялись в целостную последовательность, соответствующую правильному решению. Вероятность ее выполнения была неустойчива в пределах опыта, 76.4 % животных не могли повторить ее больше 2 - 3 раз подряд, вероятнее всего из-за дефицита внимании. Несмотря на высокий уровень мотивационной и двигательной активности, навык характеризовался высокой хаотичностью как семантической (из-за многочисленных ошибок), так и синтаксической (из-за разнообразия маршрутов), за счет чего уровень поведенческой организации был весьма низок.
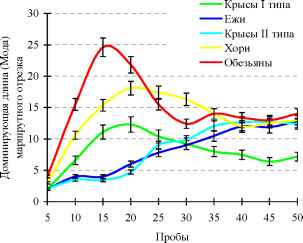
Рис. 7. Динамика ассоциативного процесса в ряду наземных млекопитающих.
Рисунок показывает положительную корреляцию между сложностью мозговой организации и скоростью ассоциативного процесса. Ежи и крысы формировали двигательную последовательность, соответствующую полному решению задачи постепенно. Хорьки, и особенно макаки, быстро сформировали различные двигательные последовательности, связывающие все семантические зоны в лабиринте, и разбивали их позже при обучении на фрагменты, соответствующие полному решению задачи.
Как показали тестовые испытания, навык, сформированный крысами I типа, был очень пластичным. В 1-ом тесте после столкновения с преградой и краткого ее обследования, крысы использовали обход уже в первом испытании, в пределах М ± SEM = 0.78 ± 0.07 мин. Второй тест показал, что интеллектуальные способности крыс I типа не выходили за рамки процедурного понимания задачи (цепной условный рефлекс), так как выполнение нового правильного решения задачи в измененной среде улучшался постепенно, отражая процесс нового обучения [Ещенко, 1998; Костенкова, Никольская, 2000]. Согласно Гальперину [1976] только "инсайтное" выполнение правильного решения задачи в новых условиях было бы доказательством того, что животное имело представление о структуре задачи. Опираясь на поведенческие и познавательные особенности, описанные выше, мы определили эту группу крыс как животные с процедурным типом мышления. Такие особенности крыс I типа как повышенная возбудимость и высокая скорость обработки информации позволяют решать проблемную ситуацию по принципу: чем выше скорость "перебора" сформированных ассоциаций, тем выше вероятность успеха.
Другую часть обучившихся крыс (59.8 ± 1.3 %) мы определили как тормозный II тип. Согласно психофизиологическим параметрам эти крысы существенно отличались от возбудимого I типа (Рис. 3 - 6). Низкий уровень двигательной активности (М ± S.E.M. = 246.5 ±
3.93 зоны/опыта; p <0.05) и более высокий уровень пассивных стрессовых реакций, чем у крыс I типа (М ± S.E.M = 0.18 ± 0.06; p <0.05), являлись типичными поведенческими особенностями, проявляемые этими крысами в начале обучения (рис. 4B). Они демонстрировали повышенный уровень осторожности к новым объектам типа дверей и кормушек, и начали брать пищу только с 3-го - 4-го опыта после того, как они полностью обследовали экспериментальную среду. Ошибочные действия, в отличие от крыс I типа, провоцировали тормозный эффект -последующее за ошибкой непродолжительное подавление или снижение двигательной активности [Ещенко, 1998; Kostenkova et al., 1999; Костенкова, Никольская, 2000]. Ошибки у крыс II типа сопровождались различными пассивными формами психо-эмоциональных проявлений типа груминг, застывание, длительностью М. ± S.E.M. = 1.8 ± 0.05 минуты (рис. 3). Процесс обучения осуществлялся на фоне низкой вероятности успеха и доминирования в поведении пассивно-оборонительных реакций, вследствие чего кривая накопления проб, эффективности поведения и организованности навыка имели логистическую динамику и напоминали инсайт-подобное обучение. Накопление различных двигательных последовательностей и их удлинение у крыс II типа, в отличие от I типа, происходило последовательно (рис. 7) и не сопровождалось явлением сенситизации. Несмотря на то, что мотивационная активность увеличивалась по экспоненте, как у крыс I типа, а организованность навыка развивались по логистическому типу, скачкообразный подъем поведенческой эффективности коррелировал с резким снижением лишних действий и ошибок (рис. 4B). После того, как процесс решения задачи был завершен, поведение крыс II типа становилось очень устойчивым и стереотипным, что нашло отражение в высоких значениях поведенческой организованности, ОН = 0.78 ± 0.05.
Таблица 2.
Показатели формирования и переделки 4-звенного навыка у наземных млекопитающих.
Первичное обучение | Переделка | навыка | |||||
Когнитивные | Когнитивные | Соотношение (Кз 2/Кз 1)x 102 | |||||
Виды | Пробы | Опыты | затраты Кз_1 | Пробы | Опыты | затраты Кз_2 | |
Ежи | 56.23 ± 2.47 | 7.84 ± 0.69 | 21.52 ± 1.05 | 25.75 ± 1.22 | 2.96 ± 0.21 | 2.21 ± 0.10 | 10.27 |
Крысы I типа | 112.33 ± 3.74** | 7.52 ± 0.77 | 55.66 ± 2.31** | 35.53 ± 2.12 | 3.33 ± 0.27 | 4.30 ± 0.51* | 7.72 |
Крысы II типа | 53.72 ± 1.21 | 8.33 ± 0.48 | 16.38 ± 0.53 | 23.84 ± 1.11 | 2.31 ± 0.14 | 1.72 ± 0.12 | 10.24 |
Хори | 45.58 ± 1.09 | 6.82 ± 0.56 | 8.87 ± 0.26** | 20.41 ± 1.40 | 2.33 ± 0.17 | 1.38 ± 0.09 | 15.55 |
Обезьяны | 33.24 ± 2.31 | 6.44 ± 1.43 | 2.87 ± 0.22* | 18.42 ± 1.78 | 2.01 ± 0.12 | 1.03 ± 0.07 | 35.89 |
Приведены Means ± SEM. * = p < 0.05 and ** = p < 0.001 (t-test Student) для групп в сравнении с крысами II типа.
Крысы II типа проигрывали I типу по своей поведенческой пластичности, показывая признаки инертности. При внезапном столкновении с препятствием, преграждающим выбранный маршрут, они предпочитали решать эту ситуацию либо «в лоб», перелезая через перегородку, чтобы достичь подкрепляемую кормушку, либо сразу же переходили на половинчатое решение: Вх - К1 - Вых - Вх. Однако, это не подразумевало, что животные не знали альтернативный маршрут, так как 67.3 % крыс II типа использовали обход после 3 - 4 " прямых" попыток. Интересно, что решение обхода не было предпочтительным, поскольку
92.8 % крыс сразу же исключали заблокированный маршрут из навыка и начинали выполнять половинчатое решения задачи, посещая доступную кормушку по самому короткому маршруту на симметричной половине лабиринта.
Такое временное изменение поведения не влияло на семантическую структуру предварительно сформированного навыка, крысы легко возвращались к прежнему целостному решению задачи после удаления барьера. Можно предположить, что выполнение половинчатого решения задачи "разумно" с точки зрения его прагматической ценности и " более экономично" для получения пищу.
Примечательно, что крысы II типа оказались более успешными в тесте по переделке информационного значения кормушек [Никольская с соавт., 1978; Ещенко, 1998; Kostenkova et al., 1999]. Динамика реорганизации поведения имела "инсайтную" кривую, познавательные затраты были значительно меньше, чем у крыс I типа (табл. 2). Вначале переделки, несмотря на изменение условий задачи, крысы II типа многократно повторяли предварительно выученную поведенческую программу в течение 8 - 14 проб, прежде чем начинали апробировать различные варианты решений, включая новое. Для большинства крыс II типа (72.2 %) эта стадия продолжалась один опыт, реже два; в этот период вероятность нового решения задачи было крайне низкой (М ± S.E.M. = 0.18 ± 0.06). Однако уже в следующем опыте (обычно 2- 3-м опыте теста) животные переходили к новому решению задачи. Быстрый переход к использованию предварительно ложных кормушек (выполнение нового решения задачи) отражает, по нашему мнению, концептуальное понимание задачи (S <-» S ассоциация, или условный рефлекс 2-ого порядка в терминах Павлова). Процесс переделки для крыс II типа состоял в перепроверке ранее сформированного правила поведения. Таким образом, если правило оставалось тем же самым, семантическая последовательность элементов задачи не менялась. На основе поведенческих и познавательных особенностей, описанных выше, мы определили эту группу крыс как животные с концептуальным типом мышления.
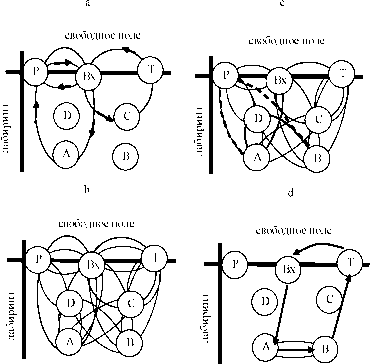
Рис. 8. Схематическое изображение поведенческих тактик на различных стадиях обучения.
Кругами показаны семантические элементы задачи, стрелками - траектория перемещения животного в лабиринте. A. Стадия 1 - тактика знакомства с новой средой, особенность которой состояло в том, что животное возвращалось в свободное поле (стартовая позиция) по траектории зеркально симметричной той, с помощью которой оно приближалось к незнакомой зоне лабиринта.
B.Стадия 2 - ненаправленный обход значимых зон лабиринта. Вероятность связи между двумя или более семантическими зонами не превышала 0.05 ±0.01 (М ± S.E.M.).
C.Стадия 3 - ситуационно - обусловленный обход; поведение животного приобретало пространственно-ориентированные черты. Животные использовали внешние ориентиры или различного рода ошибки как подсказки для перехода на симметричную половину лабиринта, чтобы получить вторую порцию пищи (жирная и пунктирная линии). D. Стадия 4 - целенаправленный обход в
содержание:
[стр.Введение] [стр.1] [стр.2] [стр.3] [стр.4] [стр.5] [стр.6] [стр.7] [стр.8] [стр.9] [стр.10] [стр.11] [стр.12] [стр.13] [стр.14] [стр.15] [стр.16] [стр.17] [стр.18] [стр.19]