 |
 Как обустроить мансарду?  Как создать искусственный водоем?  Как наладить теплоизоляцию?  Как сделать стяжку пола?  Как выбрать теплый пол?  Зачем нужны фасадные системы?  Что может получиться из балкона? |
Главная страница » Энциклопедия строителя
содержание:
[стр.Введение] [стр.1] [стр.2] [стр.3]
страница - 2
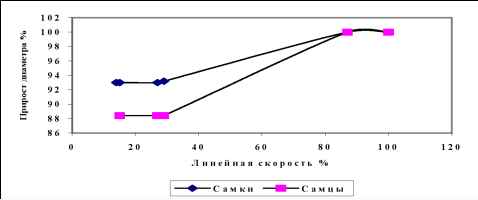
Рис. 4. Нормализованные кривые зависимости диаметра (D100) от линейной скорости потока (VL100), соответствующий полной релаксации артерии.
Пусковым стимулом эндотелий-зависимой поток-индуцируемой реакции в артериях является напряжение сдвига - сила, действующая на единицу боковой поверхности трубки со стороны движущейся в ней жидкости, прямо пропорциональная скорости движения и вязкости жидкости [4]:
ох = n*dro/dn
где ох - напряжение сдвига, п - вязкость жидкости, dco - скорость движения жидкости, dn - градиент скорости в направлении нормали движения жидкости. Из формулы следует, что при одинаковом увеличении скорости потока большее напряжение сдвига возникнет в тех сосудах, где выше значения вязкости крови. В подопытных группах выявлены различия (Р<0,05) в вязкости крови, которые составили: 3,4±0,21 спз у самок, 4±0,28 спз у самцов кроликов. Очевидно, что потоковый стимул, возникающий в сосудах самцов выше потокового стимула возникающего в сосудах самок. Поток-индуцируемый прирост диаметра артерий самцов выше на 64% прироста диаметра самок. Вязкость крови самок и самцов отличается на 17%. Таким образом, отличия в вязкости крови не более чем на 30% (в таком соотношении находятся проценты прироста диаметра и разности вязкости крови) объясняют связанные с полом различия в приросте диаметра, вызванном увеличением скорости потока. Оставшиеся 70% различий в реактивности обусловлены прочими вышеперечисленными факторами последовательности звеньев потоковой реактивности.
Из рисунка 3 А и 4 видно, что в зоне допороговой скорости потока диаметр артерий самцов ниже диаметра артерий самок. Особенностью эндотелий-зависимой поток-индуцируемой регуляции артерий является то, что в области VL100 имеет место полная релаксация сосудов. Ранее нами в исследовании биомеханических свойств артерий
показано, что диаметр дилатированного максимальным потоком сосуда оказывается равным диаметру этого же сосуда обработанного высокими дозами папаверина [17]. Поскольку механизм поток-зависимой регуляции способен полностью исчерпывать возможности сосуда к релаксации [17], величина потоковой реакции (aD%) 11,58% у самцов и 7,05 % у самок отражает степень релаксации сосудов от исходного диаметра в отсутствие потокового стимула (D0). В диапазоне скоростей от V0 до VL100 различия хода кривых зависимости диаметра от линейной скорости определяются эндотелий-зависимым поток-индуцируемым механизмом релаксации гладких мышц, поэтому меньшие значения диаметра артерий самцов в зоне допороговой скорости свидетельствуют о том, что в сосудах самцов имеет место более высокое напряжение гладкой мускулатуры, или другими словами этим сосудам свойственен более высокий тонус. Эту величину мы обозначили как Т0 - тонус в отсутствие потокового стимула, Т0 сосудов самцов выше нулевого тонуса сосудов самок.
Обсуждение результатов
Определенные суждения о причине половых различий в тонусе сонных артерий можно вынести на основании данных, полученных на коронарных артериях [15]. Экспериментальные, инструментальные, клинико-биохимические исследования свидетельствуют о способности эстрогенов при различных путях введения вызывать снижение тонуса как интактных, так и предварительно спазмированных вазоконстрикторами коронарных артерий [28]. Эстрогены могут быстро оказывать влияние на тонус сосудов путем прямого воздействия на электрические свойства биомембран и трансмембранную ионную проницаемость [3]. Снижение тонуса коронарных сосудов в ответ на стероидные и нестероидные эстрогены не зависит от адренэргических, холинэргических, гистаминэргических механизмов, а связано с блокадой трансмембранного транспорта ионов Са2+ [35]. В настоящее время обнаружены две изоформы рецепторов эстрогенов (ERU и ERp) [19, 27], причем ER участвует в реализации большинства физиологических эффектов, оказываемых эстрадиолом. Функции ERp остаются пока невыясненными [19]. Согласно многим данным, 170-эстрадиол значительно повышает уровень секреции КО клетками эндотелия сосудов [40], что служит еще одним фактором вышеописанного вазодилатирующего эффекта эстрадиола.
Эти факты экстраполированные на сонные артерии могут объяснить почему сосуды самок имеют более низкий тонус и меньший угол наклона кривых зависимости диаметра
от линейной скорости несмотря на более низкий порог чувствительности к изменению скорости потока.
Как показано в работе Maddox с соавт. [30] эндотелий самок и самцов аорты крыс отличается по чувствительности к простагаландину F2a. Эндотелий аорты самцов имеет более высокую чувствительность к PgF2a, в этой же работе не было обнаружено различий в сократимости сосудистых гладких мышц. Обработка тестостероном повышала чувствительность эндотелия сосудов самок крыс к PgF2a, но не влияла на сократимость гладкой мускулатуры.
Таким образом эстрогены могут выступать в роли модулятора состояния гладких мышц и быть ответственны за различия в исходном тонусе (Т0), поток-зависимом приросте диаметра на единицу скорости (dD/dVL) в целом у самок и самцов кроликов.
Выявленные половые различия в реакциях сонных артерий на увеличение скорости потока только на 30% определяются отличиями в вязкости крови, и как следствием, различным потоковым стимулом в сосудах самок и самцов кроликов, ведущим фактором является отличающийся тонус исследованных сосудов, что может быть обусловлено половыми особенностями гормонального статуса экспериментальных животных.
Литература
1.Балашов С. А., Мелькумянц A. M. Изменение тонуса сонных артерий кошек в ответ на изменение скорости кровотока // Бюллетень экспериментальной биологии и медицины. - 1984. - N 11. - С. 515-517.
2.Захарова Г. Н. Облитерирующий эндартериит конечностей. - Саратов: Изд-во Саратовского ун-та, 1973. - 252 с.
3.Кобрин В. И., Маноах М., Варон Д. и др. Влияние 17-Р эстрадиолсульфата на трансмембранные потенциалы кардиомиоцитов сердца морской свинки // Бюллетень экспериментальной биологии и медицины. - 1996. - № 4. - С. 370 - 373.
4.Коган В. Б. Теоретические основы типовых процессов химической технологии. -Л., 1977.
5.Корж А. Н. Современные представления о структуре, функции и биологической роли сосудистого эндотелия // Междунар. мед. ж. (Харьков). - 2003. - Т. 9, № 1. -С. 130—134.
6.Манухина Е. А, Малышев И. В., Микоян Н. Н. Увеличение продукции оксида азота в органах крысы при тепловом шоке // Бюллетень экспериментальной биологии и медицины. - 1996, №5. - С.520-524.
содержание:
[стр.Введение] [стр.1] [стр.2] [стр.3]